Oxygen isotopic evidence from the deep oceans, as well as geomorphological and radiometric evidence from Iceland suggest that northern hemispheric ice sheets formed much later than the Antarctic ice sheets. Northern hemisphere ice sheets may have started to form in the late Miocene, about 6 million years ago, but they grew to reach sea-level all around the Arctic Ocean only by about 3.1 million years ago. A major increase in ice-volume occurred about 2.75 to 2.45 million years ago. For the last 2.45 million years large ice sheets on the Northern hemisphere waxed and waned cyclically, with a larger amplitude from 0.9 Ma on, at the specific Milankovich orbital frequencies. There has been much interest in the process of development of northern hemisphere ice sheets during the last 2.5 million years: this period is so recent that many organisms alive at the beginning of the formation of these ice sheets, or their close relatives, are still living, so that we know their environmental preferences. Deposits formed during this most recent period are at the surface in many regions, and thus can be studied in land exposures, and from the ocean floor in short (<10m long) piston cores, which can be taken fairly easily from non-specialized research vessels.
Land studies had demonstrated that there had been more than one 'ice age', a period during which large ice masses covered the northwest European and North American continents. The number of such periods, glaciations, was thought to be four or five (on the European mainland: Donau, Günz, Mindel, Riss, Würm). From studies of ocean cores, however, it became readily apparent that many more fluctuations in extent of the ice sheets had occurred: about 22 fluctuations were recognized over the last 800,000 years, up to 65 over the last 2 million yr. The extent to which ice-sheets reached southward varied from period to period, so that not each "ice age" was as extreme; many "partial ice ages" occurred These fluctuations were documented first by studies of calcareous nannofossils and planktonic foraminifera in the northern Atlantic and Pacific Oceans.
In the present oceans these planktonic microfloras/faunas occur in roughly latitudinal assemblages (modified by surface current systems). We can thus analyze the foraminiferal assemblage as we find them in the top of cores (i.e., the foraminifera as we find them in the sediment, reflecting present-day circumstances). We can then compare the faunal assemblages as we find them in the core tops with the temperature of the surface waters above the core sites. We can also compare the faunas in core tops with older faunas from samples lower in the cores. In the past the boundaries between the faunal assemblages have been at different latitudes from the present position, and the faunal boundaries, and thus presumably the boundaries between surface water masses of varying temperatures, have been moving north and south continuously. More quantitative work was needed to make estimates of the magnitudes of these environmental changes.
The next step was to translate the information from faunal assemblages (so many percent of species A, so many percent of species B, and so on, through about 29 species) into an estimate of the temperature of the surface waters for present day faunas for which water temperatures can be measured. We do this by submitting the paleontological census data (relative abundance data, % of each species) to a multivariate analysis (i.e., a statistical analysis using many variables at the same time). We then derive new parameters which consist of some combination of the old parameters (species percentages), and then relate the multivariate, derived parameters to observed oceanographic data (mainly sea surface temperature, SST). We derive a set of equations called transfer functions in which faunal percentages can be expressed as sea-surface temperature. We can derive both summer and winter temperatures, because different species reflect coldest and warmest months temperatures. A transfer function is thus a set of equations relating oceanographic parameters to faunal data (more in general: in a transfer function parameters are expressed as parameters with other dimensions). We can, for instance, use information on diatom populations in lake to calculate the acidity of lake waters, and then use fossil diatom data to reconstruct the history of pH of lakes.
Using transfer functions, we can thus derive estimates for surface water temperatures in the past, and we can derive a set of temperatures for winter and summer. These records show very strong fluctuations over time, with the magnitude of the fluctuations varying by latitude: the largest fluctuations were found at the middle-latitudes (40 to 60 oN).
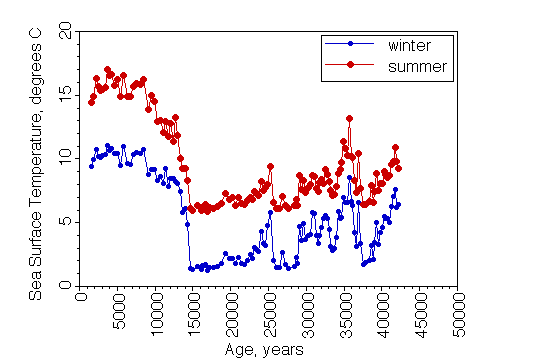
Shown here is a temperature reconstruction for a core taken at about 50oN in the North Atlantic, using data on the abundance of 20 species of planktonic foraminifera. Note the rapid end of the ice age just after 15,000 years ago, and the large changes in ocean surface temperatures at this mid latitude location.
The fluctuations in faunal content were mirrored in global fluctuations in the oxygen isotopic values values of benthic and planktonic foraminifera. The oxygen isotopic records of planktonic foraminifera show an overall global difference between glacial and non-glacial periods of about 1.9o/oo (heavier during cold periods). Early interpretations were that this record was mainly the result of temperature fluctuations of about 6-7 oC even in the tropics (about 1.4o/oo temperature, 0.5 o/oo ice volume effect). Comparison of benthic and planktonic records, however, demonstrated that oxygen isotope fluctuations in both are of similar magnitude. At locations where the temperature of bottom waters is now about 0.5 oC, however, the deep water temperature cannot have been 6 to 7 oC lower during a glacial (that would mean frozen deep oceans). Thus there must be a large ice volume effect in both planktonic and benthic records. If the temperature change in the bottom waters was just a few degrees, than that in the surface waters at low latitudes must have been similar. This conclusion was supported by transfer function analysis of planktonic foraminiferal faunas from low latitudes, where the temperature of surface waters apparently fluctuated by only a few degrees C at most (1.5 + 1.5oC).
More recently, however, the issue
of how much tropical
temperatures changed during
glacials has become hotly debated once again. A new method of
measuring paleotemperatures uses the ratios of the concentrations of
Mg to Ca or Sr to Ca in various shells, including
corals. It turns out that this ratio is temperature-dependent,
and larger at higher temperatures. The specific temperatures found
for several corals in the tropics suggest that the
tropic
coastal waters were cooler by at least 5 to 6 oC
during glacials. It remains
the question in how such large temperature changes also occurred in
open oceans: temperatures always change much more on continents than
in open ocean.
We can check the inference that
open ocean tropical temperatures changed little (i.e., by a few
degrees at maximum) by using estimates
of sea-level lowering. If the
sea level was about 130 m lower during maximum glacial development
(best estimate from raised coral-reef terraces, dated with
14C and 230Th/234U, various methods
of radiometric age determmination), then we can estimate what the
difference in ice volume to cause this lowering of sea
level would have caused in the global oxygen isotopic values. If
we assume an average
oxygen isotopic composition of the ice
sheet of &endash;35
o/oo, then the
maximum
change in average isotopic composition of the oceans as a result
of melting enough ice to cause a sea level rise of 130 m was about
1.3
o/oo:
10 m sea level change corresponds to about 0.1 o/oo
d
18O (heavier during
the last glacial; i.e., a
large part of the 1.9 o/oo
signal)
The fluctuations in sea surface
temperatures and ice volume are cyclical, with a periodicity agreeing
to that in astronomical parameters influencing the amount of sunlight
(insolation) received at different locations on Earth, the
Milankovich
frequencies (see handout
on Milankovich).
Remember that these periodicities are:
Carbon isotopic records also show changes between glacial and interglacial time periods. These records show a depletion oby about 0.8 o/oo in benthic values during glacials (thus lighter values of total dissolved inorganic carbon during glacials); the benthic values can be seen as representing the average of the deep global ocean. This was interpreted originally as resulting from less storage of organic material in the terrestrial biosphere during glacials. The land biomass was supposedly smaller, because large, now forested areas (before human deforestation) were barren tundras during glacials. The shallow water shelves were dry during glacial periods (lower sea level), thus organic-rich sediments on the shelves could be eroded and their light organic carbon added to the oceans. This idea, however, led to the prediction that there would have been more CO2 in the atmosphere during glacials (about 150 ppm), because less carbon would have been locked in the biosphere and shallow water sediment.
This prediction was incorrect: measurements of CO2 concentration in bubbles of air trapped in polar ice demonstrated that there was a lower concentration of CO2 in the atmosphere during glacials (by about 90 ppm, leading to a glacial CO2 concentration of about 200ppm). The contradiction could be partially solved by comparing benthic and planktonic carbon isotopic records. The planktonic records show much less change between glacials and interglacials than the benthic records, so that the difference in d13C values in surface and deep waters (called Dd13C) was larger (about 2.1) during glacials than during interglacials (about 1.5). This suggests that productivity in the oceans was higher during glacials. Why would this be?
At the start of a glacial, sea level drops as a result of storage of water in ice caps. Thus the organic matter on shelves is eroded into the oceans, thus leading to decreased values of d13C in Total Dissolved Carbon in the oceans. This erosion, however, adds not only carbon, but also nutrients from the oxidized organic material, leading to increased nutrient contents in ocean water. The increased amount of nutrients (fertilizer) may then lead to increased oceanic primary productivity.
The temperature differences from high to low latitudes in atmosphere and ocean increased during glacial periods, because there was strong cooling at high latitudes, little or no cooling at low latitudes. These increased gradients may have led to stronger, more vigorous currents and winds, leading to more vigorous upwelling, which could lead to even higher surface ocean productivity. It has also been proposed that the ratio of N and P to C in organic matter (the Redfield ratio) might have differed during glacial times, which simplified means, that organisms might have been able to produce more organic material with less nutrients (N and P).
The effect of decreasing productivity during interglacials is to decrease Dd13C (the difference in carbon isotopic values for planktonic and benthic foramifera, i.e., the difference in carbon isotopic values of dissolved carbonate in surface and bottom waters), and bring the benthic and planktonic records closer together, as observed. But one would expect the planktonic curve to show the largest excursion (because that's where the change in productivity occurs), while the benthic values stayed almost unchanged, because the deep ocean is so much larger than the surface ocean. But the benthic values fluctuated most, while the planktonic values showed slight fluctuations only. This suggest that there was not only a productivity effect, but also a reservoir effect, resulting from decreased areas of forests during glacials - leading to lighter isotopic values in total dissolved carbon during glacials. For the planktonic foraminifera the effects of productivity and reservoir almost cancel each other out: during glacials there is higher productivity, leading to heavier values of total dissolved carbon and thus of the planktonic tests, but at the same time light carbon is being added to the oceans from the terrestrial biosphere reservoir, leading to lighter values of the total dissolved carbon. Presently, about 0.4 o/oo of the changes in benthic foraminiferal d13C values is attributed to changes in terrestrial biosphere, about 0.4 o/oo to oceanic productivity changes.
This is not the last complication
of the
d13C
records, especially for the records from the North Atlantic. In the
present ocean, deep
water is formed in the North
Atlantic, leading to the
presence of "young", well-oxygenated, 13C-enriched water
in the northern Atlantic, and the existence of
strong
d13C
differences between deep waters in Atlantic and Pacific. This
difference became much smaller or disappeared during glacials.
Benthic foraminiferal faunal evidence suggests that faunas living in
the north Atlantic during glacials were below much less oxygen-rich
waters in that area. This combined evidence suggests that
deep-water formation in the north Atlantic stopped or diminished
strongly during glacials (see class readings by Broecker). In the
North Atlantic records we thus see combined effects of
productivity
changes,
circulation
changes, and
reservoir
changes. The difference
between glacial and interglacial benthic foraminiferal values in the
North Atlantic is thus larger than in the Pacific.
To get back
(finally) to the CO2 levels of the
atmosphere: as a result of
this increased
surface ocean productivity
the surface waters (where productivity finds place) are more
strongly depleted in total dissolved carbon (TDC) during glacials
than during interglacials. These surface waters (NOT the deep
waters), however, equilibrate with the atmosphere, leading to
decreased CO2 contents of the atmosphere (the atmosphere
"sees" only the carbon-depleted surface waters, not the
average-ocean).
So how about
Milankovich
cycles
causing ice ages?
The description of glacials-interglacials as resulting from cyclical changes of insolation as a result of orbital changes is thus essentially probably correct, but much too simplified.
This suggests that the insolation changes are the trigger, and that subsequent feedback reinforces the climate signal.
The last problem is that the character of the glacial-interglacial
fluctuations has changed. From about 2.5 m.y. to about 0.9 m.y. the
41,000 years periodicity dominated the total signal, after that the
100,000 year periodicity was more dominant, and the amplitude of the
glacial-interglacial changes increased. This change in character has
not yet been fully explained, but some investigators supposed that
the Pliocene uplift of the Himalayan Mountains might have influenced
global weather-patterns (especially the position of the
upper-atmospheric jet streams), leading to more extreme swings in
climate.
This change in deep-water formation in the North Atlantic must also have influenced the world's climate, because the North Atlantic water (before sinking) cools strongly, and thus gives off heat to the north Atlantic environment. Lack of this deep water formation may thus have helped cool the northeastern Atlantic during glacials.
The processes of "switching on and off" glacials are thus very complex, and the complexity is increasing rapidly with more and more knowledge. A more detailed knowledge of the way in which the ocean-atmosphere system works, and of the role of the biosphere in shaping climate (by its effects on the carbon cycle) is needed before we can begin to understand even this latest, best-known part of Earth history.