|
Dana Royer Assistant Professor Department of Earth and Environmental Sciences Wesleyan University Exley Science Center 445 (265 Church St.) Middletown, CT 06459-0139 office: 860.685.2836; lab: 860.685.2873; fax: 860.685.3651 droyer@wesleyan.edu |
||||||||
|
After
nearly 90 years of use, the analysis of leaf shape remains the most reliable
means to reconstruct terrestrial paleotemperatures from before the Pleistocene.
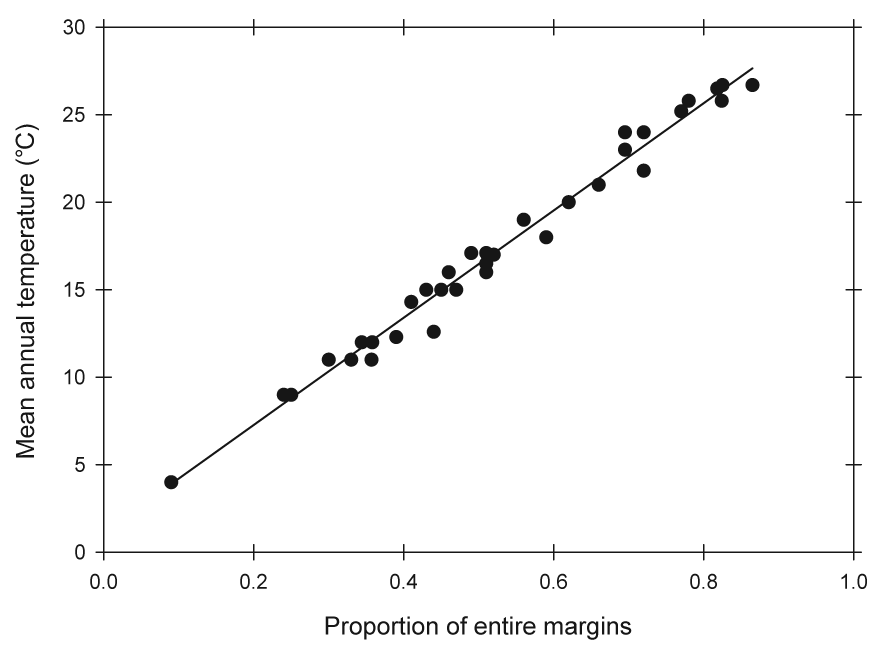
In particular, the strong correlation observed in living forests between
the proportion of plant species that have untoothed leaf margins and mean
annual temperature is widely applied to fossil floras ("leaf-margin analysis"; see Figure 1). Given
the importance of leaf-margin analysis, it is striking that both the mechanistic underpinnings
behind the correlation are poorly known and that potentially more accurate
methods based on leaf shape are not considered reproducible. We are addressing
these gaps in understanding. First, we have tested the hypothesis that teeth enhance plant
growth at the beginning of the growing season when temperatures are limiting.
Testing involved tracking seasonal patterns
of leaf-margin gas exchange of a warm and cold temperate flora, enabling
quantitative comparisons among species as well as groups of species.
Significant results are: physiological activity at leaf margins is greatest early in the
growing season; toothed margins are more active than untoothed margins; and leaf margins
are more active in species native to colder region. Thus, teeth increase transpiration and
photosynthate production early in the growing season, maximizing carbon gain when temperature
is limiting. This helps explain the core
observation underlying most paleoclimate estimates based on fossil leaf teeth.
Second, we have developed a new method for quantifying the shapes of leaves based on computerized image analysis. Relative to leaf-margin analysis, this method describes more fully the sizes and shapes of leaves. The analysis of 17 test sites from eastern North and Central America shows that many size and shape variables correlate significantly with mean annual temperature, indicating a coordinated, convergrent evolutionary response of fewer teeth, smaller tooth area, and lower degree of blade dissection in warmer environments. Multiple linear regressions based on a subset of shape variables (tooth area / leaf area; leaf perimeter / perimeter after teeth are removed; floral proportion of toothed species) produce more accurate estimates of mean annual temperature than leaf-margin analysis. This opens the exciting possibility for reconstructing ancient paleotemperatures from fossil leaves more accurately than ever before. Finally, we are exploring correlations between the shapes of leaves and leaf economic variables. Leaf economic variables describe how fast or slow a plant "spends" its nutrient resources, and include leaf photosynthetic rate, leaf nitrogen content, leaf mass per area, and leaf lifespan. These variables tell you something important about the functioning of a given ecosystem (i.e., how fast or slow it is turning over), but unfortunately they cannot be directly quantified on fossil plants. However, we are finding significant correlations between leaf economic and tooth area variables, suggesting that it may be possible to reconstruct paleo-leaf economics via proxy. Our ultimate goal is to reconstruct, from single outcrops, climate and leaf economics from the shapes of fossil leaves. These data could be coupled with other outcrop data such as insect herbivory, leaf stomatal index, and the stable carbon isotopic composition of fossil leaf organic material, providing powerful snapshots of ancient ecosystems.
Related publications [pdf] Royer DL, Sack L, Wilf P, Lusk CH, Jordan GJ, Niinemets Ü, Wright IJ, Westoby M, Cariglino B, Coley PD, Cutter AD, Johnson KR, Labandeira CC, Moles AT, Palmer MB, Valladares F. 2007. Fossil leaf economics quantified: calibration, Eocene case study, and implications. Paleobiology, 33: 574-589. The leaves of most modern vascular plants show an inverse correlation between stomatal index (the percentage of leaf epidermal cells that are stomata) and the partial pressure of atmospheric CO2. By using phylogenetically conservative taxa such as Ginkgo (maidenhair tree) or Metasequoia (dawn redwood), these modern relationships can be applied to the fossil record to reconstruct paleo-CO2. The early Paleogene (c. 65 to 45 Ma) and middle Miocene (17.5 to 15.5 Ma) represent the warmest periods in Earth’s history since the demise of the dinosaurs. They therefore represent ideal intervals to study the dynamics of Earth’s climate system during a globally warm phase. Recent reconstructions of CO2 for the early Paleogene range from < 300 ppm to > 3000 ppm, indicating gross inconsistencies, while reconstructions for the middle Miocene are all < 350 ppm, suggesting that CO2 and global temperatures were not as tightly coupled as they are today. Using a series of 25 fossil plant localities containing cuticle of Ginkgo and Metasequoia, we have been able to reconstruct CO2 for these two intervals. Our results indicate that CO2 was between 300 and 450 ppm during these two periods. These data represent the first quantitative stomatal-based CO2 reconstructions for these two intervals, and have helped to refine our understanding of CO2 during these globally warm times. Our data also bear on the issue of present-day climate change, as it is possible that CO2 may approach levels in the near future that have not been surpassed for the last 65 million years. Related publications [pdf] Royer DL. 2003. Estimating latest Cretaceous and Tertiary atmospheric CO2 from stomatal indices. In: Wing SL, Gingerich PD, Schmitz B, Thomas E (eds). Causes and Consequences of Globally Warm Climates in the Early Paleogene. Geological Society of America Special Paper, 369: 79-93. [pdf] Beerling DJ, Lomax BH, Royer DL, Upchurch GR, Kump LR. 2002. An atmospheric pCO2 reconstruction across the Cretaceous-Tertiary boundary from leaf megafossils. Proceedings of the National Academy of Sciences USA, 99: 7836-7840. [pdf] Beerling DJ, Royer DL. 2002. Fossil plants as indicators of the Phanerozoic global carbon cycle. Annual Review of Earth and Planetary Sciences, 30: 527-556. [pdf] Beerling DJ, Royer DL. 2002. Reading a CO2 signal from fossil stomata. New Phytologist, 153: 387-397. [pdf] Royer DL. 2002. Estimating latest Cretaceous and Tertiary PCO2 from stomatal indices [Ph.D. thesis]. Yale University, New Haven, 163 p. [pdf] Royer DL, Berner RA, Beerling DJ. 2001. Phanerozoic atmospheric CO2 change: Evaluating geochemical and paleobiological approaches. Earth-Science Reviews, 54: 349-392. [pdf] Royer DL, Wing SL, Beerling DJ, Jolley DW, Koch PL, Hickey LH, Berner RA. 2001. Paleobotanical evidence for near present day levels of atmospheric CO2 during part of the Tertiary. Science, 292: 2310-2313. [pdf] Royer DL. 2001. Stomatal density and stomatal index as indicators of paleoatmospheric CO2 concentration. Review of Palaeobotany and Palynology, 114: 1-28. |